Chapter 13: Data requirements for biological effects studies - which components and attributes of the marine biota to examine?
- 13.1 Components
- 13.2 Plankton and fish
- 13.3 Macrobenthos and meiobenthos
- 13.4 Hard-bottom epifauna and hard-bottom motile fauna
- 13.5 Attributes and recommendations
13.1 Components
The biological effects of pollutants can be studied on assemblages of a wide variety of marine organisms:
Pelagos
- plankton (both phytoplankton and zooplankton)
- fish (pelagic and demersal)
Benthos (soft-bottom)
- macrobenthos
- meiobenthos
- microbenthos, not much used in community studies
Benthos (hard-bottom)
- epifauna (encrusting forms, e.g. corals)
- motile fauna (both macrofauna and meiofauna in e.g. algae, holdfasts and epifauna)
These various components of the biota each have certain practical and conceptual advantages and disadvantages for use in biological effects studies. These are discussed in this chapter, and an example is given for each of the components (although not all of these examples are directly concerned with pollution effects).
13.2 Plankton and fish
Plankton
The advantages of plankton are that:
a) Long tows over relatively large distances result in community samples which reflect integrated ecological conditions over large areas. They are therefore useful in monitoring more global changes.
b) Identification of macro-planktonic organisms is moderately easy, because of the ready availability of appropriate literature.
The disadvantage of plankton is that, because the water masses in which they are suspended are continually mobile, they are not useful for monitoring the local effects of a particular pollutant source.
Example: Continuous Plankton Recorder
Plankton samples have been collected from ‘ships of opportunity’ plying their usual commercial routes across the NE Atlantic since the late 1940s (e.g. Colebrook (1986) ) and continue today, their historical continuity (through the Sir Alister Hardy Foundation for Ocean Science, Plymouth, UK) giving the survey ever greater importance with respect to climate change monitoring. The CPR plankton recorders collect samples through a small aperture, and these are trapped on a continuously winding roll of silk so that each section of silk contains an integrated sample from a relatively large area. This has enabled long term trends in plankton abundance to be assessed; e.g. Colebrook (1986) describes a gradual decline in both zooplankton and phytoplankton since the early 1950s, with an upturn in the 1980s (Fig. 13.1).
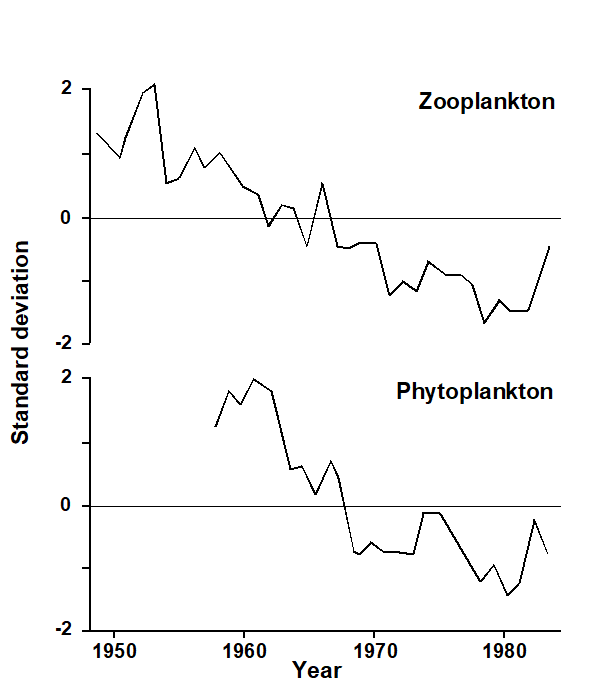
Fig. 13.1. Continuous Plankton Recorder Survey of the NE Atlantic {p}. First principal components for zooplankton and phytoplankton over the first 35 years of the survey (from
Colebrook (1986)
). Graphs scaled to zero mean and unit variance.
Fish
The advantages of fish are that:
a) Because of their mobility they are again more useful for studying general rather than local effects, but some demersal fish communities may show site fidelity, such as the coral-reef fish in the example below.
b) The taxonomy of fish is relatively easy, in all parts of the world.
c) Fish are of immediate commercial and public interest, and so studies of fish communities are more directly related to the needs of environmental managers than, for example, the meiobenthos (despite the fact, of course, that the latter are vitally important to the early life-stages of fish!).
The disadvantages of fish are that:
a) Strictly quantitative sampling which is equally representative of all the species in the community is difficult. The overall catching efficiency of nets, traps etc. is often unknown, as are the differing abilities of species to evade capture or their susceptibility to be attracted to traps. Visual census methods are also not free from bias, since some species will be more conspicuous in colouration or behaviour than other dull secretive species.
b) Uncertainty about site fidelity is usually, but not always, a problem.
Example: Maldives coral reef-fish
For a study in the Maldive islands, Dawson-Shepherd, Warwick, Clarke et al. (1992) used visual census methods to compare reef-fish assemblages at 23 coral reef-flat sites, 11 of which had been subjected to coral mining for the construction industry and 12 were non-mined controls. The MDS (Fig. 13.2) clearly distinguished mined from non-mined sites.
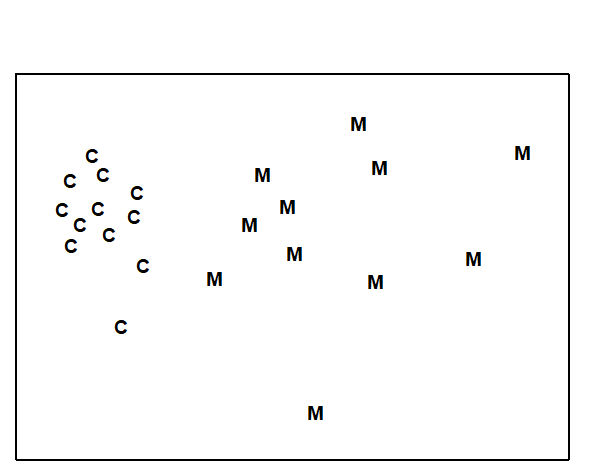
Fig. 13.2. Maldive Islands, coral-reef fish {M}. MDS ordination of fish species abundance data from mined (M) and control (C) reef-flats (stress = 0.08).
13.3 Macrobenthos and meiobenthos
Macrobenthos
The advantages of soft-bottom macrobenthos are that:
a) They are relatively non-mobile and are therefore useful for studying the local effects of pollutants.
b) Their taxonomy is relatively easy.
c) Quantitative sampling is relatively easy.
d) There is an extensive research literature on the effects of pollution, particularly organic enrichment, on macrobenthic communities, against which specific case-histories can be evaluated.
This combination of advantages has resulted in the soft-bottom macrobenthos being probably the most widely used component of the marine biota in environmental impact studies. Despite this, they do have several disadvantages:
a) Relatively large-volume sediment samples must be collected, so that sampling requires relatively large research ships.
b) Because it is generally not practicable to bring large volumes of sediment back to the laboratory for processing, sieving must be done at sea and is rather labour intensive and time consuming (therefore expensive).
c) The potential response time of the macrobenthos to a pollution event is slow. Their generation times are measured in years, so that although losses of species due to pollution may take immediate effect, the colonisation of new species which may take advantage of the changed conditions is slow. Thus, the full establishment of a community characterising the new environmental conditions may take several years.
d) The macrobenthos are generally unsuitable for causality experiments in mesocosms, because such experiments can rarely be run long enough for fully representative community changes to occur, and recruitment of species to mesocosm systems is often a problem because of their planktonic larval stages (see Chapter 12).
Example: Amoco Cadiz oil-spill
The sensitivity of macrobenthic community structure to pollution events, when using multivariate methods of data analysis, is discussed in Chapter 14. The response of the macrobenthos in the Bay of Morlaix to the Amoco Cadiz oil-spill some 40 km away, already seen in Chapters 5, 7 and 10, is a good example of this (Fig. 13.3).
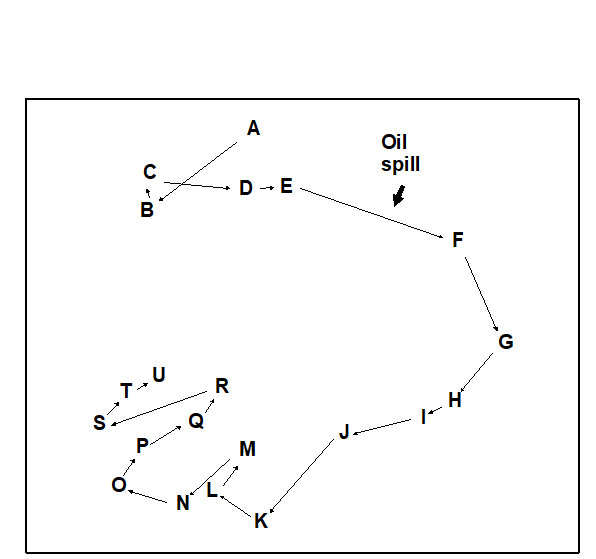
Fig. 13.3. Amoco-Cadiz oil spill, Bay of Morlaix {A}. MDS for macrobenthos at station ‘Pierre Noire’, at approximately three-monthly sampling intervals (stress = 0.09).
Meiobenthos
Apart from sharing the advantage of non-mobility, and therefore usefulness for local effects studies, the relative advantages and disadvantages of the meiobenthos are exactly the reverse of the macrobenthos. Their advantages are:
a) Because of their small size and high density in marine sediments, quantitative sampling of the meiobenthos is easy from small ships, open boats etc.
b) The small volume of the samples means that they can easily be transported to the laboratory, and need not be processed on board ship.
c) Their generation times are usually measured in months rather than years, so that their potential response time to pollution events is much faster than that of the macrobenthos.
d) Because of this fast response time, and direct benthic rather than planktonic development, the meiobenthos are good candidates for causality experiments in experimental microcosms and mesocosms.
The disadvantages of meiobenthos are that:
a) Their taxonomy is considered difficult. Identification of almost all the meiobenthic taxa to species level presents difficulties even in Europe and N America, and in many parts of the world the fauna is almost completely unknown. However, to a considerable degree, three factors mitigate against this problem:
i. The robustness of community analyses to the use of taxonomic levels higher than species (see Chapter 10).
ii. The cosmopolitan nature of most meiobenthic genera.
iii. The increasing availability of easily used keys to meiobenthic genera. For example, the pictorial keys to marine nematodes of Platt & Warwick (1988) have been used successfully worldwide.
b) Community responses of the meiobenthos to pollution are not as well documented as for the macrobenthos, and there is only a modest body of information in the literature against which particular case-histories can be evaluated.
Example: Soldier crab disturbance of nematode assemblages, Tasmania
This natural field experiment was first met in Chapters 6 and 12. It will be remembered that the nematode diversity profiles were affected by the crab disturbance (Fig. 12.2), whereas no significant effect was noted for copepods (Fig. 13.4). Many nematode species are more sedentary in habit than copepods, often adhering to sand-grains by secretions from their caudal glands, and some species prefer conditions of low oxygen concentration or are obligate anaerobes. The so called ‘thiobiotic’ meiofaunal community contains many nematode species, but apparently no copepods. Non-bioturbated sediments will have a vertical gradient in physical and chemical conditions ranging from wave-disturbed sediments with an oxiphilic meiofauna community near the surface to a stable sediment with a thiobiotic community deeper down. Dramatic disturbance by crabs, of the kind found at this site, will inevitably destroy this gradient, so that the whole sediment column will be well aerated and unstable. This reduction in habitat complexity is probably the most parsimonious explanation for the reduction in nematode species diversity.
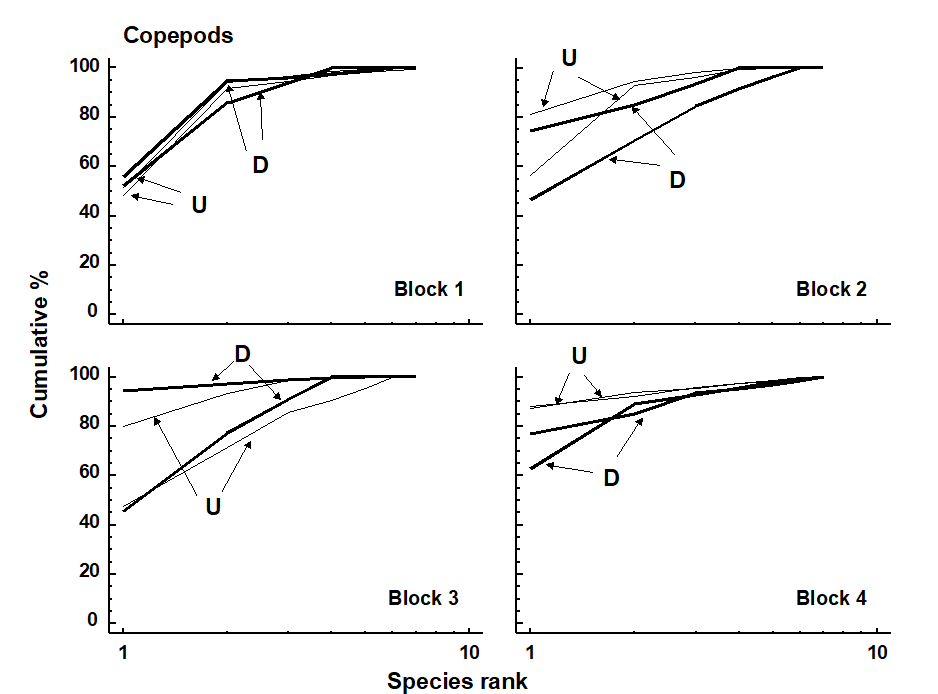
Fig. 13.4. Tasmania, Eaglehawk Neck {T}. k-dominance curves for disturbed (D) and undisturbed (U) copepod samples in in 4 separate sampling blocks.
The differential response of these two components of the meiobenthos has been elaborated here in order to demonstrate how a knowledge of the biology of these components can aid in the interpretation of community responses to perturbation. The macrobenthos and meiobenthos may also respond differently to different kinds of perturbation (e.g. physical disturbance, pollution) so that a comparative study of both may be indicative of the cause.
Example: Macrobenthos and meiobenthos in Hamilton Harbour, Bermuda
Fig. 13.5 shows the average k-dominance curves for the macrobenthos and the nematode component of the meiobenthos at six stations in Hamilton Harbour. For the macrobenthos, the curves at three of the stations (H3, H4 & H6) are much more elevated than the other three, suggesting some kind of perturbation at these sites. For the nematodes, however, all curves are closely coincident. There must therefore be some form of perturbation affecting the macrobenthos but not the meiobenthos, and it was suggested by Warwick, Platt, Clarke et al. (1990) that this is more likely to be physical disturbance of the sediment resulting from the regular passage of large cruise liners within the harbour, rather than pollution. This is because the macrobenthos are much more dependent on sediment stability to maintain diversity than are the meiobenthos.
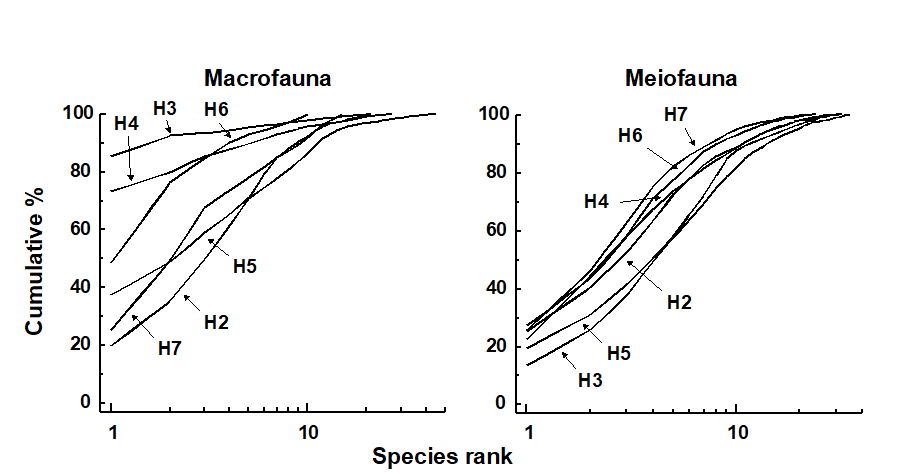
Fig. 13.5. Hamilton Harbour, Bermuda {H}. k-dominance curves for macrobenthos (left) and meiobenthic nematodes (right) at six stations (H2-H7).
13.4 Hard-bottom epifauna and hard-bottom motile fauna
Hard-bottom epifauna
The advantages of using hard-bottom encrusting faunas, reef-corals etc. are:
a) They are immobile and therefore good for local effects studies.
b) A major advantage over sedimentary faunas is that non-destructive (visual/photographic) sampling is possible.
The disadvantages are:
a) Remote sampling is more difficult. Intertidal or shallow subtidal sites can be surveyed (the latter by divers); remote cameras require a greater level of technical sophistication but great strides have been made with this technology in recent decades.
b) Enumeration of colonial organisms is difficult, so that abundance units such as number of colonies or percentage cover must be used – this only becomes a problem when it is necessary to combine data on colonial organisms with that on motile species (see page 5.9). However, biomass measurements are difficult to make.
Example: Indonesian reef corals
The example shown in Fig. 13.6, of the effects of the 1982-3 El Niño on reef coral communities at South Pari Island, was seen in Chapter 10. A clear difference is seen in community composition between 1981 and 1983, with a more steady pattern of change thereafter, though without full reversion to the initial state.

Fig. 13.6. Indonesian reef-corals {I}. MDS for coral species percentage cover data for South Pari Island (10 replicate transects in each year). 1=1981, 3=1983 etc. (stress = 0.25).
Hard-bottom motile fauna
The motile fauna living on rocky substrates and associated with algae, holdfasts, hydroids etc. has rarely been used in pollution impact studies because of its many disadvantages:
a) Remote sampling is difficult.
b) Quantitative extraction from the substrate, and comparative quantification of abundances between different substrate types, are difficult.
c) Responses to perturbation are largely unknown.
d) A suitable habitat (e.g. algae) is not always available. A solution to this problem, and also problem (b),
that has sometimes been tried in practice, is to deploy standardised artificial substrates, e.g. plastic mesh pan-scrubbers, along suspected pollution gradients in the field, allowing these to become colonised.
Example: Metazoan fauna of intertidal seaweed samples from the Isles of Scilly
The entire metazoan fauna (macrofauna + meiofauna) was examined from five species of intertidal macro-algae (Chondrus, Laurencia, Lomentaria, Cladophora, Polysiphonia) each collected at eight sites near low water from rocky shores on the Isles of Scilly, UK ( Gee & Warwick (1994a) , Gee & Warwick (1994b) ). The MDS plots for meiobenthos and macrobenthos were very similar, with the algal species showing very similar relationships to each other in terms of their meiofaunal and macrofaunal community structure (Fig. 13.7). The structure of the weed therefore clearly influenced community structure in both these components of the benthic fauna.
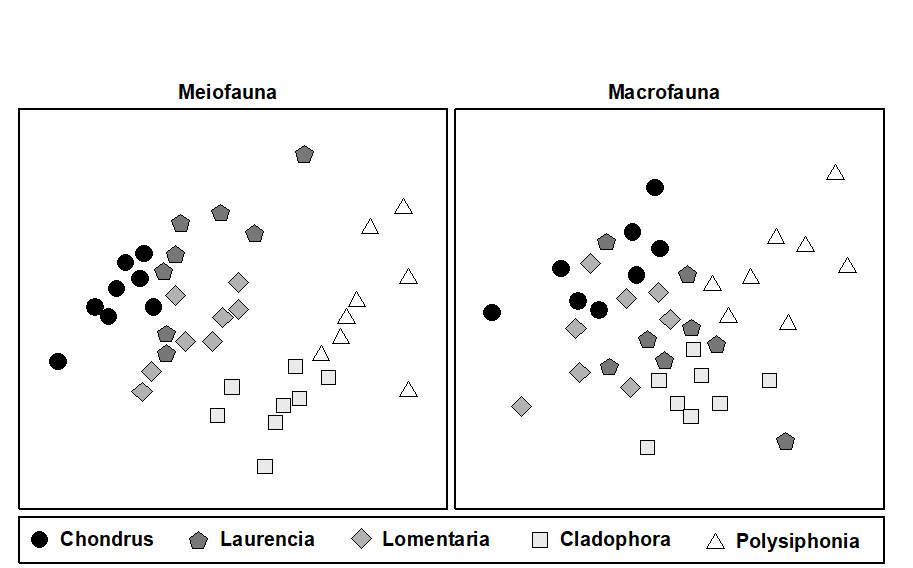
Fig. 13.7. Scilly Isles seaweed fauna {S}. MDS of standardised $\sqrt{} \sqrt{}$-transformed meiofauna and macrofauna species abundance data. The five seaweed species are indicated by different symbol and shading conventions (stress = 0.19, 0.18).
13.5 Attributes and recommendations
Attributes
Species abundance data are by far the most commonly used in environmental impact studies at the community level. However, the abundance of a species is perhaps the least ecologically relevant measure of its relative importance in a community, and we have already seen in Chapter 10 that higher taxonomic levels than species may be sufficient for environmental impact analyses. So, when planning a survey, consideration should be given not only to the number of stations and number of replicates to be sampled, but also to the level of taxonomic discrimination which will be used, and which measure(s) of the relative importance of these taxa will be made.
Abundance, biomass and production
As a measure of the relative ecological importance for soft-sediment and water-column sampling of species, biomass is better than abundance, and production in turn is better than biomass. However, the determination of annual production of all species within a community over a number of sites or times would be so time consuming as to be completely impracticable.¶ We are therefore left with the alternatives of studying abundances, biomasses, or both. Abundances are marginally easier to measure, biomass may be a better reflection of ecological importance, and measurement of both abundance and biomass opens the possibility of comparing species-by-sites matrices based on these two different measures (e.g. by the ABC method discussed in Chapter 8).
In practice, multivariate analyses of abundance and biomass data often give remarkably similar results, despite that fact that the species mainly responsible for discriminating between stations are usually different. In Fig. 13.8, for example, the Frierfjord macrobenthos MDS configurations for abundance and biomass are very similar but it is small polychaete species which are mainly responsible for discriminating between sites on the basis of abundance, and species such as the large echinoid Echinocardium cordatum which discriminate the sites on the basis of biomass.
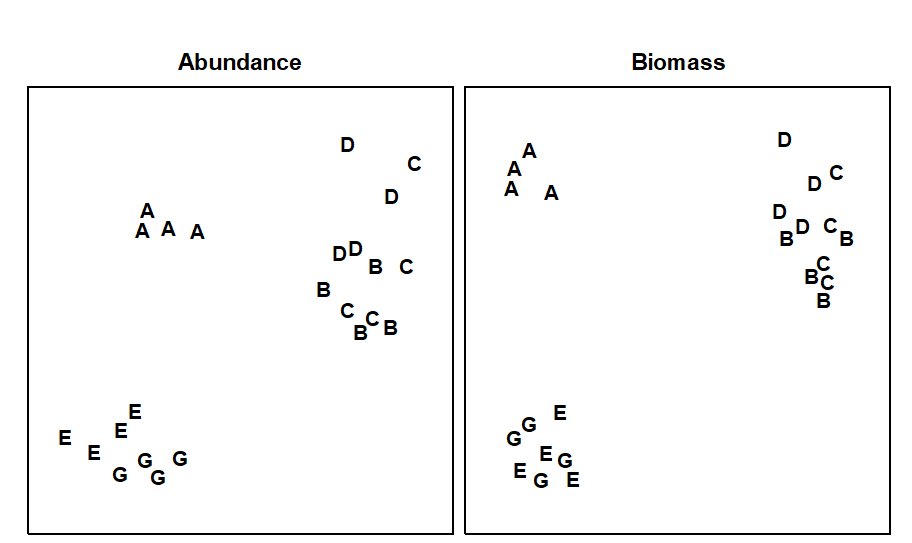
Fig. 13.8. Frierfjord macrofauna {F}. MDS ordinations for abundance and biomass of the 4 replicates at each of the 6 sites (stress = 0.10, 0.08).
Species or higher taxa
We have already seen in Chapter 10 that, in many pollution-impact studies, it has been found for both graphical and multivariate analyses that there is surprisingly little loss of information when the species data are aggregated to higher taxa, e.g. genera, families or sometimes even phyla. For the detection of pollution impact, initial collection of data at the level of higher taxa would result in a considerable saving of time (and cost) in the analysis of samples. Such a strategy would, of course, be quite inappropriate if the objective were to be differently defined, for example, the quantification of biodiversity properties.
Recommendations
It is difficult to give firm recommendations as to which components or attributes of the biota should be studied, since this depends on the problem in hand and the expertise and funds available. In general, however, the wider the variety of components and attributes studied, the easier the results will be to interpret. A broad approach at the level of higher taxa is often preferable to a painstakingly detailed analysis of species abundances. If only one component of the fauna is to be studied, then consideration should be given to working up a larger number of stations/replicates at the level of higher taxa in preference to a small number of stations at the species level. Of course, a large number of replicated stations at which both abundance and biomass are determined at the species level is always the ideal!
¶ Although relative ‘production’ of species can be approximated using empirical relationships between biomass, abundance and production, and these ‘production’ matrices subjected to multivariate analysis, see Chapter 15.