Chapter 4: Ordination of samples by principal components analysis (PCA)
- 4.1 Ordinations
- 4.2 Principal components analysis
- 4.3 Example: Garroch Head macrofauna
- 4.4 PCA for environmental data
- 4.5 Example: Dosing experiment, Solbergstrand mesocosm
4.1 Ordinations
An ordination is a map of the samples, usually in two or three dimensions, in which the placement of samples, rather than representing their location in space (or time), reflects the similarity of their biological communities. To be more precise, distances between samples on the ordination attempt to match the corresponding dissimilarities in community structure: nearby points have very similar communities, samples which are far apart have few species in common or the same species at very different levels of abundance (or biomass). The word ‘attempt’ is important here since there is no uniquely defined way in which this can be achieved. Indeed, when a large number of species fluctuate in abundance in response to a wide variety of environmental variables, with many species being affected in different ways, the community structure is essentially high-dimensional and it may be impossible to obtain a useful two or three-dimensional representation.
So, as with cluster analysis, several methods have been proposed, each using different forms of the original data and varying in their technique for approximating high-dimensional information in low-dimensional plots. They include:
a) Principal Components Analysis, PCA (see, for example, Chatfield & Collins (1980) );
b) Principal Co-ordinates Analysis, PCO ( Gower (1966) );
c) Correspondence Analysis and Detrended Correspondence Analysis, DECORANA ( Hill & Gauch (1980) );
d) Multi-Dimensional Scaling, MDS; in particular non-metric MDS (see, for example, Kruskal & Wish (1978) ).
A comprehensive survey of ordination methods is outside the scope of this manual. As with clustering methods, detailed explanation is given only of the techniques required for the analysis strategy adopted throughout the manual. This is not to deny the validity of other methods but simply to affirm the importance of applying, with understanding, one or two techniques of proven utility. The two ordination methods selected are therefore (arguably) the simplest of the various options, at least in concept.
a) PCA is the longest-established method, though the relative inflexibility of its definition limits its practical usefulness more to multivariate analysis of environmental data rather than species abundances or biomass; nonetheless it is still widely encountered and is of fundamental importance.
b) Non-metric MDS does not have quite such a long history (though the key paper, by Kruskal, is from 1964!). Its clever and subtle algorithm, some years ahead of its time, could have been contemplated only in an era in which significant computational power was foreseen (it was scarcely practical at its time of inception, making Kruskal’s achievement even more remarkable). However, its rationale can be very simply described and understood, and many would argue that the need to make few (if any) assumptions about the data make it the most widely applicable and effective method available.
4.2 Principal components analysis
The starting point for PCA is the original data matrix rather than a derived similarity matrix (though there is an implicit dissimilarity matrix underlying PCA, that of Euclidean distance). The data array is thought of as defining the positions of samples in relation to axes representing the full set of species, one axis for each species. This is the very important concept introduced in Chapter 2, following equation (2.13). Typically, there are many species so the samples are points in a very high-dimensional space.
A simple 2-dimensional example
It helps to visualise the process by again considering an (artificial) example in which there are only two species (and nine samples).
| Sample | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Abundance | Sp.1 | 6 | 0 | 5 | 7 | 11 | 10 | 15 | 18 | 14 |
| Sp.2 | 2 | 0 | 8 | 6 | 6 | 10 | 8 | 14 | 14 |
The nine samples are therefore points in two dimensions, and labelling these points with the sample number gives:
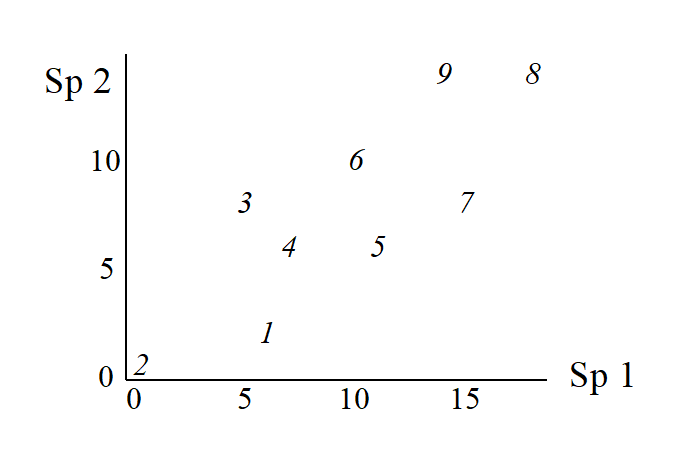
This is an ordination already, of 2-dimensional data on a 2-dimensional map, and it summarises pictorially all the relationships between the samples, without needing to discard any information at all. However, suppose for the sake of example that a 1-dimensional ordination is required, in which the original data is reduced to a genuine ordering of samples along a line. How do we best place the samples in order? One possibility (though a rather poor one!) is simply to ignore altogether the counts for one of the species, say Species 2. The Species 1 axis then automatically gives the 1-dimensional ordination (Sp.1 counts are again labelled by sample number):
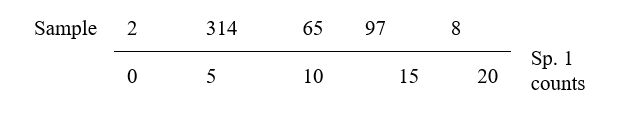
(Think of this as projecting the points in the 2-dimensional space down onto the Sp.1 axis). Not surprisingly, this is a rather inaccurate 1-dimensional summary of the sample relationships in the full 2-dimensional data, e.g. samples 7 and 9 are rather too close together, certain samples seem to be in the wrong order (9 should be closer to 8 than 7 is, 1 should be closer to 2 than 3 is) etc. More intuitively obvious would be to choose the 1-dimensional picture as the (perpendicular) projection of points onto the line of ‘best fit’ in the 2-dimensional plot.
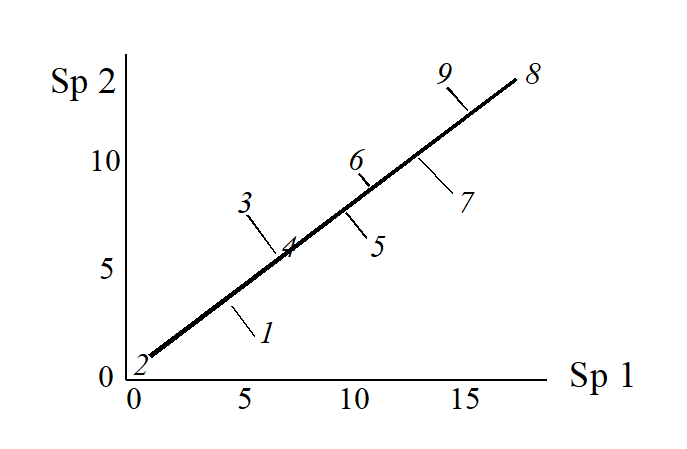
The 1-dimensional ordination, called the first principal component axis (PC1), is then:
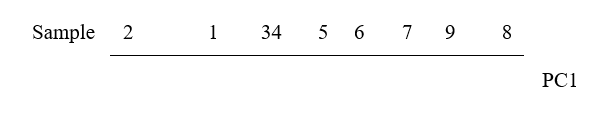
and this picture is a much more realistic approximation to the 2-dimenensional sample relationships (e.g. 1 is now closer to 2 than 3 is, 7, 9 and 8 are more equally spaced and in the ‘right’ sequence etc).
The second principal component axis (PC2) is defined as the axis perpendicular to PC1, and a full principal component analysis then consists simply of a rotation of the original 2-dimensional plot:
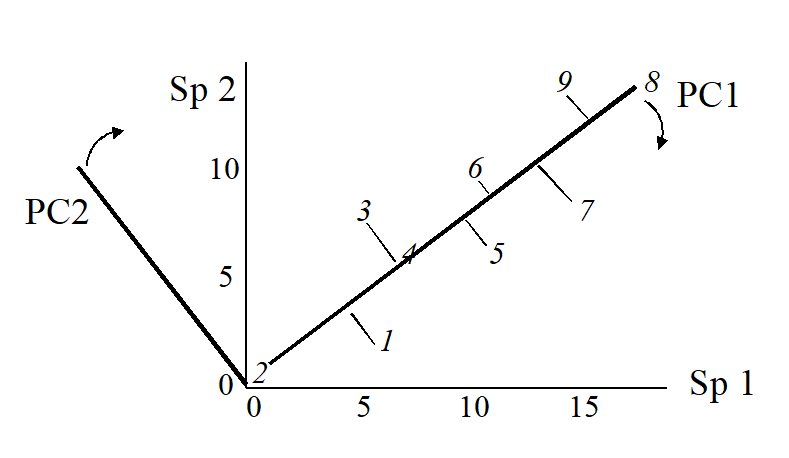
to give the following principal component plot.
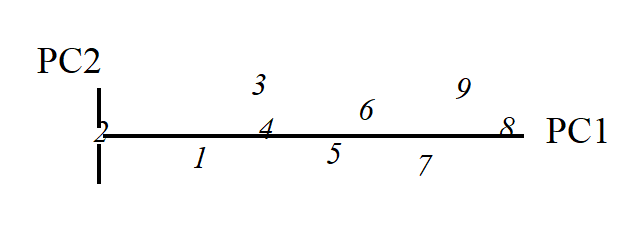
Obviously the (PC1, PC2) plot contains exactly the same information as the original (Sp.1, Sp.2) graph. The whole point of the procedure though is that, as in the current example, we may be able to dispense with the second principal component (PC2): the points in the (PC1, PC2) space are projected onto the PC1 axis and relatively little information about the sample relationships is lost in this reduction of dimensionality.
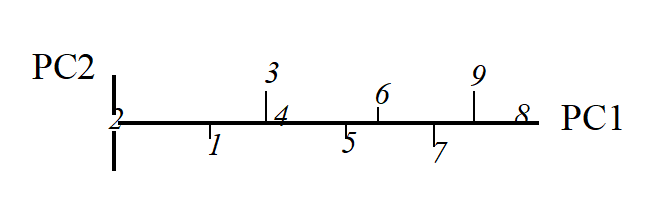
Definition of PC1 axis
Up to now we have been rather vague about what is meant by the ‘best fitting’ line through the sample points in 2-dimensional species space. There are two natural definitions. The first chooses the PC1 axis as the line which minimises the sum of squared perpendicular distances of the points from the line.¶ The second approach comes from noting in the above example that the biggest differences between samples take place along the PC1 axis, with relatively small changes in the PC2 direction. The PC1 axis is therefore defined as that direction in which the variance of sample points projected perpendicularly onto the axis is maximised. In fact, these two separate definitions of the PC1 axis turn out to be totally equivalent† and one can use whichever concept is easier to visualise.
Extensions to 3-dimensional data
Suppose that the simple example above is extended to the following matrix of counts for three species.
| Sample | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Abundance | Sp.1 | 6 | 0 | 5 | 7 | 11 | 10 | 15 | 18 | 14 |
| Sp.2 | 2 | 0 | 8 | 6 | 6 | 10 | 8 | 14 | 14 | |
| Sp.3 | 3 | 1 | 6 | 6 | 9 | 11 | 10 | 16 | 15 |
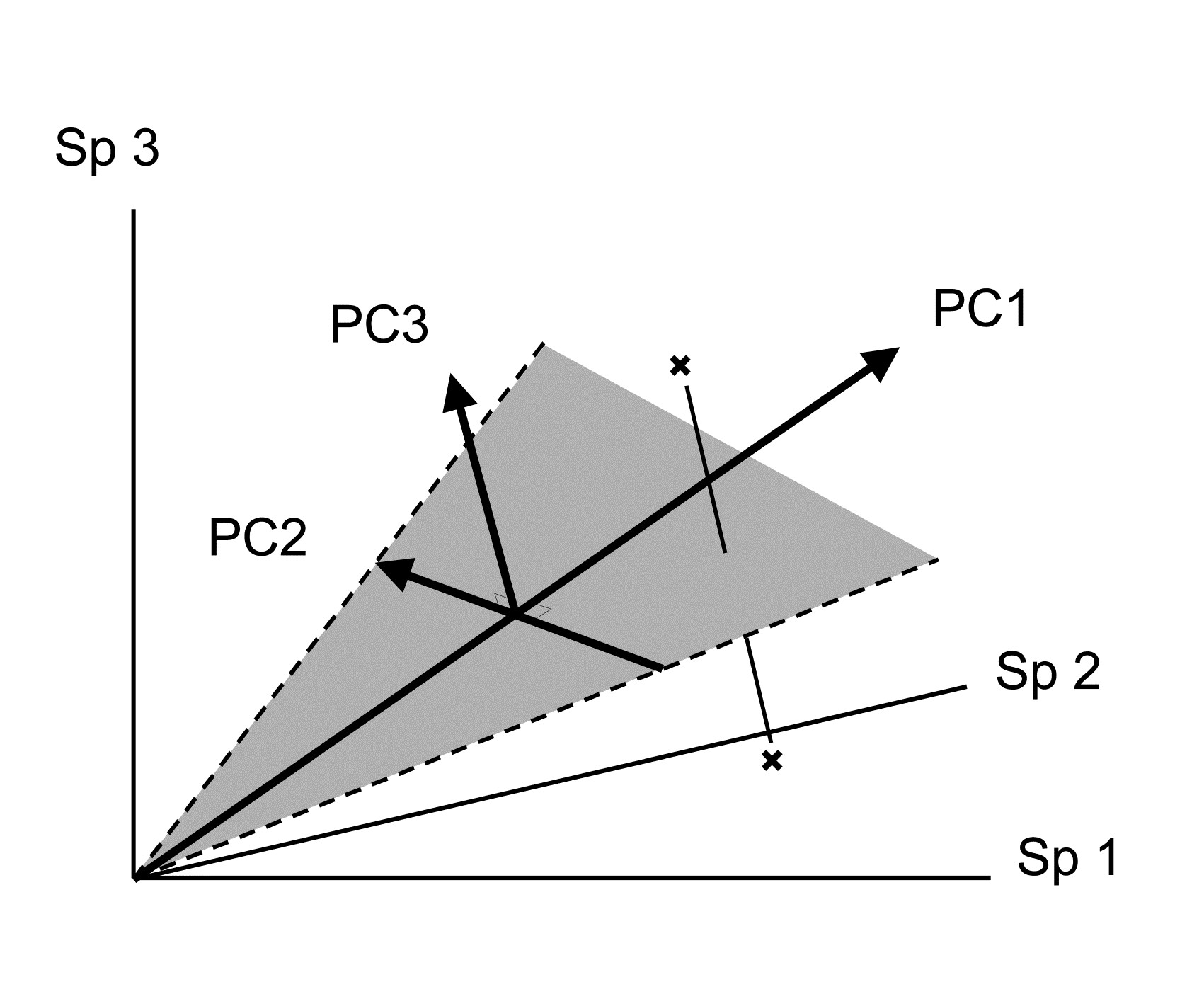
Samples are now points in three dimensions (Sp.1, Sp.2 and Sp.3 axes) and there are therefore three principal component axes, again simply a rotation of the three species axes. The definition of the (PC1, PC2, PC3) axes generalises the 2-dimensional case in a natural way:
PC1 is the axis which maximises the variance of points projected perpendicularly onto it;
PC2 is constrained to be perpendicular to PC1, but is then again chosen as the direction in which the variance of points projected perpendicularly onto it is maximised;
PC3 is the axis perpendicular to both PC1 and PC2 (there is no choice remaining here).
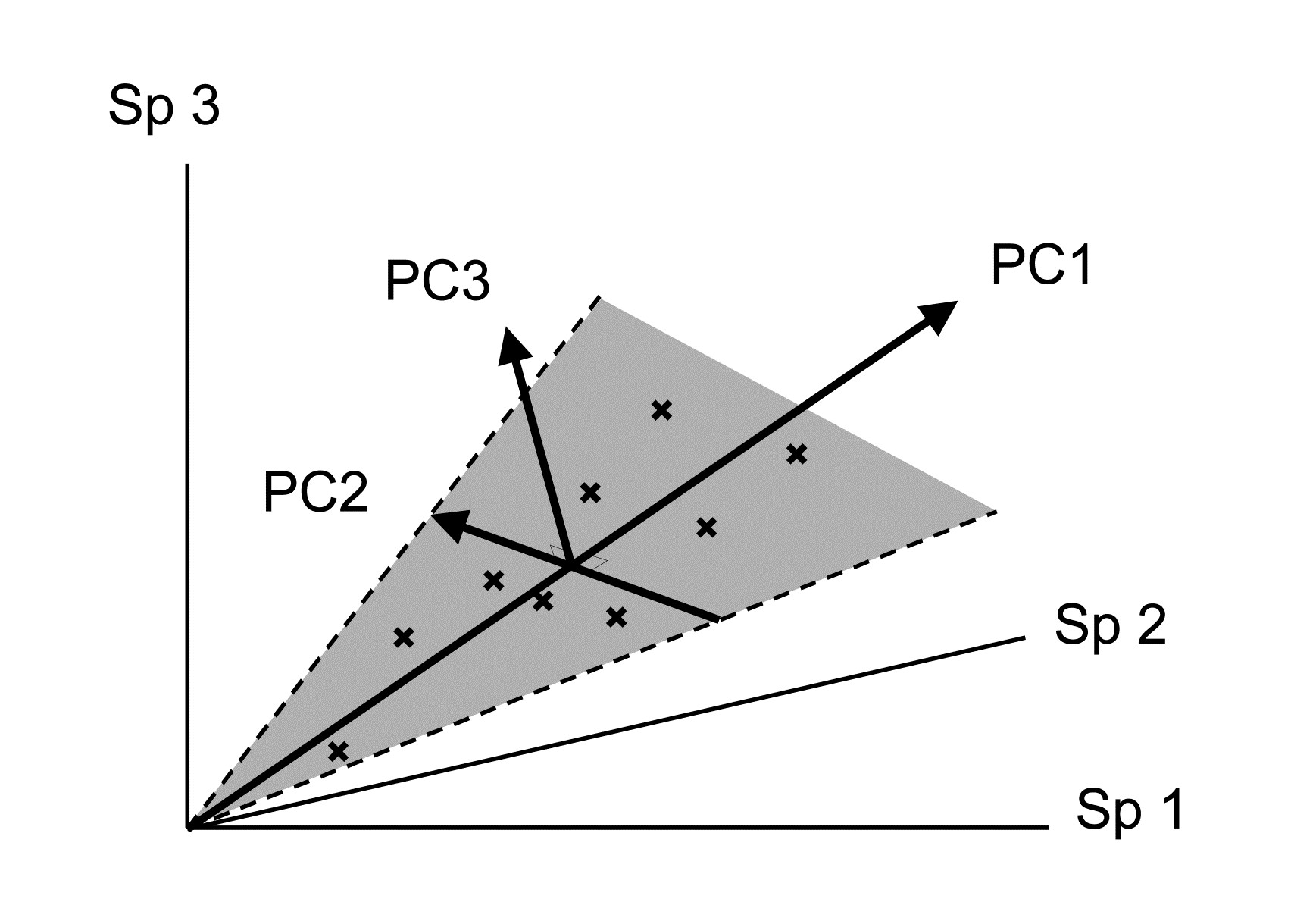
An equivalent way of visualising this is again in terms of ‘best fit’: PC1 is the best fitting line to the sample points and, together, the PC1 and PC2 axes define a plane (grey in the above diagram) which is the best fitting plane.
Algebraic definition
The above geometric formulation can be expressed algebraically. The three new variables (PCs) are just linear combinations of the old variables (species), such that PC1, PC2 and PC3 are uncorrelated. In the above example:
$$ PC1 = 0.62 \times Sp.1 + 0.52 \times Sp.2 + 0.58 \times Sp.3 $$ $$ PC2 = –0.73 \times Sp.1 + 0.65 \times Sp.2 + 0.20 \times Sp.3 \tag{4.1} $$ $$ PC3 = 0.28 \times Sp.1 + 0.55 \times Sp.2 – 0.79 \times Sp.3 $$
The principal components are therefore interpretable (in theory) in terms of the counts for each original species axis. Thus PC1 is a sum of roughly equal (and positive) contributions from each of the species; it is essentially ordering the samples from low to high total abundance. At a more subtle level, for samples with the same total abundance, PC2 then mainly distinguishes relatively high counts of Sp.2 (and low Sp.1) from low Sp.2 (and high Sp.1); Sp.3 values do not feature strongly in PC2 because the corresponding coefficient is small. Similarly the PC3 axis mainly contrasts Sp.3 and Sp.2 counts.
Variance explained by each PC
The definition of principal components given above is in terms of successively maximising the variance of sample points projected along each axis, with the variance therefore decreasing from PC1 to PC2 to PC3. It is thus natural to quote the values of these variances (in relation to their total) as a measure of the amount of information contained in each axis. And the total of the variances along all PC axes equals the total variance of points projected successively onto each of the original species axes, total variance being unchanged under a simple rotation. That is, letting var(PCi) denote variance of samples on the ith PC axis and var(Sp.i) denote variance of points on the ith species axis (i = 1, 2, 3):
$$ \sum_i var ( PCi ) = \sum_i var ( Sp.i) \tag{4.2} $$
Thus, the relative variation of points along the ith PC axis (as a percentage of the total), namely
$$ P_i = 100 \frac{ var ( PCi ) } { \sum_i var ( PCi ) } = 100 \frac{ var ( PCi ) } { \sum_i var ( Sp.i ) } \tag{4.3} $$
has a useful interpretation as the % of the original total variance explained by the ith PC. For the simple 3-dimensional example above, PC1 explains 93%, PC2 explains 6% and PC3 only 1% of the variance in the original samples.
Ordination plane
This brings us back finally to the reason for rotating the original three species axes to three new principal component axes. The first two PCs represent a plane of ‘best fit’, encompassing the maximum amount of variation in the sample points. The % variance explained by PC3 may be small and we can dispense with this third axis, projecting all points perpendicularly onto the (PC1, PC2) plane to give the 2-dimensional ordination plane that we have been seeking. For the above example this is:


and it is almost a perfect 2-dimensional summary of the 3-dimensional data, since PC1 and PC2 account for 99% of the total variation. In effect, the points lie on a plane (in fact, nearly on a line!) in the original species space, so it is no surprise to find that this PCA ordination differs negligibly from that for the initial 2-species example: the counts added for the third species were highly correlated with those for the first two species.
Higher-dimensional data
Of course there are many more species than three in a normal species by samples array, let us say 50, but the approach to defining principal components and an ordination plane is the same. Samples are now points in (say) a 50-dimensional species space§ and the best fitting 2-dimensional plane is found and samples projected onto it to give the 2-dimensional PCA ordination. The full set of PC axes are the perpendicular directions in this high-dimensional space along which the variances of the points are (successively) maximised. The degree to which a 2-dimensional PCA succeeds in representing the information in the full space is seen in the percentage of total variance explained by the first two principal components. Often PC1 and PC2 may not explain more than 40-50% of the total variation, and a 2-dimensional PCA ordination then gives an inadequate and potentially misleading picture of the relationship between the samples. A 3-dimensional sample ordination, using the first three PC axes, may give a fuller picture or it may be necessary to invoke PC4, PC5 etc. before a reasonable percentage of the total variation is encompassed. Guidelines for an acceptable level of ‘% variance explained’ are difficult to set, since they depend on the objectives of the study, the number of species and samples etc., but an empirical rule-of-thumb might be that a picture which accounts for as much as 70-75% of the original variation is likely to describe the overall structure rather well.
The geometric concepts of fitting planes and projecting points in high-dimensional space are not ones that most people are comfortable with (!) so it is important to realise that, algebraically, the central ideas are no more complex than in three dimensions. Equations like (4.1) simply extend to p principal components, each a linear function of the p species counts. The ‘perpendicularity’ (orthogonality) of the principal component axes is reflected in the zero values for all sums of cross-products of coefficients (and this is what defines the PCs as statistically uncorrelated with each other), e.g. for equation (4.1):
$$ (0.62) \times (-0.73) + (0.52) \times (0.65) + (0.58) \times (0.20) = 0 $$ $$ (0.62) \times (0.28) + (0.52) \times (0.55) + (0.58) \times (-0.79) = 0 $$ $$ etc $$
The coefficients are also scaled so that their sum of squares adds to one – an axis only defines a direction not a length so this (arbitrarily) scales the values, i.e.
$$ (0.62)^2 + (0.52)^2 + (0.58)^2 = 1$$ $$ (-0.73)^2 + (0.65)^2 + (0.20)^2 = 1$$ $$etc$$
There is clearly no difficulty in extending such relations to 4, 5 or any number of coefficients.
The algebraic construction of coefficients satisfying these conditions but also defining which perpendicular directions maximise variation of the samples in the species space, is outside the scope of this manual. It involves calculating eigenvalues and eigenvectors of a p by p matrix, see
Chatfield & Collins (1980)
, for example. (Note that a knowledge of matrix algebra is essential to understanding this construction). The advice to the reader is to hang on to the geometric picture: all the essential ideas of PCA are present in visualising the construction of a 2-dimensional ordination plot from a 3-dimensional species space.
(Non-)applicability of PCA to species data
The historical background to PCA is one of multivariate normal models for the individual variables, i.e. individual species abundances being normally distributed, each defined by a mean and symmetric variability around that mean, with dependence among species determined by correlation coefficients, which are measures of linearly increasing or decreasing relationships. Though transformation can reduce the right-skewness of typical species abundance/biomass distributions they can do little about the dominance of zero values (absence of most species in most of the samples). Worse still, classical multivariate methods require the parameters of these models (the means, variances and correlations) to be estimated from the entries in the data matrix. But for the Garroch Head macrofaunal biomass data introduced on page 1.6, which is typical of much community data, there are p=84 species and only n=12 samples. Thus, even fitting a single multivariate normal distribution to these 12 samples requires estimation of 84 means, 84 variances and $_{84} C _ 2 = 84 \times 83 / 2 = 3486$ correlations! It is, of course, impossible to estimate over 3500 parameters from a matrix with only $12 \times 86 = 1032$ entries, and herein lies much of the difficulty of applying classical testing techniques which rely on normality, such as MANOVA, Fisher’s linear discriminant analysis, multivariate multiple regression etc, to typical species matrices.
Whilst some significance tests associated with PCA also require normality (e.g. sphericity tests for how many eigenvalues can be considered significantly greater than zero, attempting to establish the ‘true dimensionality’ of the data), as it has just been simply outlined, PCA has a sensible rationale outside multi-normal modelling and can be more widely applied. However, it will always work best with data which are closest to that model. E.g. right skewness will produce outliers which will always be given an inordinate weight in determining the direction of the main PC axes, because the failure of an axis to pass through those points will lead to large residuals, and these will dominate the sum of squared residuals that is being minimised. Also, the implicit way in which dissimilarity between samples is assessed is simply Euclidean distance, which we shall see now (and again much later when dissimilarity measures are examined in more detail in Chapter 16) is a poor descriptor of dissimilarity for species communities. This is primarily because Euclidean distance pays no special attention to the role that zeros play in defining the presence/absence structure of a community. In fact, PCA is most often used on variables which have been normalised (subtracting the mean and dividing by the standard deviation, for each variable), leading to what is termed correlation-based PCA (as opposed to covariance-based PCA, when non-normalised data is submitted to PCA). After normalising, the zeros are replaced by different (largish) negative values for each species, and the concept of zero representing absence has disappeared. Even if normalisation is avoided, Euclidean distance (and thus PCA) is what is termed ‘invariant to a location change’ applied throughout the data matrix, whereas biological sense dictates that this should not be the case, if it is to be a useful method for species data. (Add 10 to all the counts in Table 2.1 and ask yourself whether it now carries the same biological meaning. To Euclidean distance nothing has changed; to an ecologist the data are telling you a very different story!)
Another historical difficulty with applying PCA to community matrices was computational issues with eigen-analyses on matrices with large numbers of variables, especially when there is parameter indeterminacy in the solution, from matrices having a greater number of species than samples (p > n). However, modern computing power has long since banished such issues, and very quick and efficient algorithms can now generate a PCA solution (with n-1 non-zero eigenvalues in the p > n case), so that it is not necessary, for example, to arbitrarily reduce the number of species to p < n before entering PCA.
¶ This idea may be familiar from ordinary linear regression, except that this is formulated asymmetrically: regression of y on x minimises the sum of squared vertical distances of points from the line. Here x and y are symmetric and could be plotted on either axis.
† The explanation for this is straightforward. As is about to be seen in (4.2), the total variance of the data, var(Sp1) + var (Sp2), is preserved under any rotation of the (perpendicular) axes, so it equals the sum of the variances along the two PC axes, var(PC1) + var(PC2). If the rotation is chosen to maximise var(PC1), and var(PC1) + var(PC2) is fixed (the total variance) then var(PC2) must be minimised. But what is var(PC2)? It is simply the sum of squares of the PC2 values round about their mean (divided by a constant), in other words, the sum of squares of the perpendicular projections of each point on to the PC1 axis. But minimising this sums of squares is just the definition given of ‘best fitting line’.
§ If there are, say, only 25 samples then all 50 dimensions are not necessary to exactly represent the distances among the 25 samples – 24 dimensions will do (any two points fit on a line, any three points fit in a plane, any four points in 3-d space, etc). But a 2-d representation still has to approximate a 24-d picture!
4.3 Example: Garroch Head macrofauna
Fig. 4.1 shows the result of applying PCA to square-root transformed macrofaunal biomass data from the 65 species¶ found in subtidal sediments at 12 sites (1-12) along an E-W transect in the Firth of Clyde, Scotland ({G}, map at Fig. 1.5). A central site of the transect (site 6) is an accumulating sewage-sludge dump-ground and is subject to strong impacts of organic enrichment and heavy metal concentrations.
It makes sense to transform the biomass values, for much the same reasons as for the cluster analyses of Chapter 3, so that analysis is not dominated by large biomass values from a small number of species; here a mild square-root transform was adequate to avoid the PCA becoming over-dependent on a few outliers. There is also no need to normalise species variables: they are on comparable and meaningful measurement scales (of biomass), so PCA will naturally give more weight to species with larger (transformed) biomass.
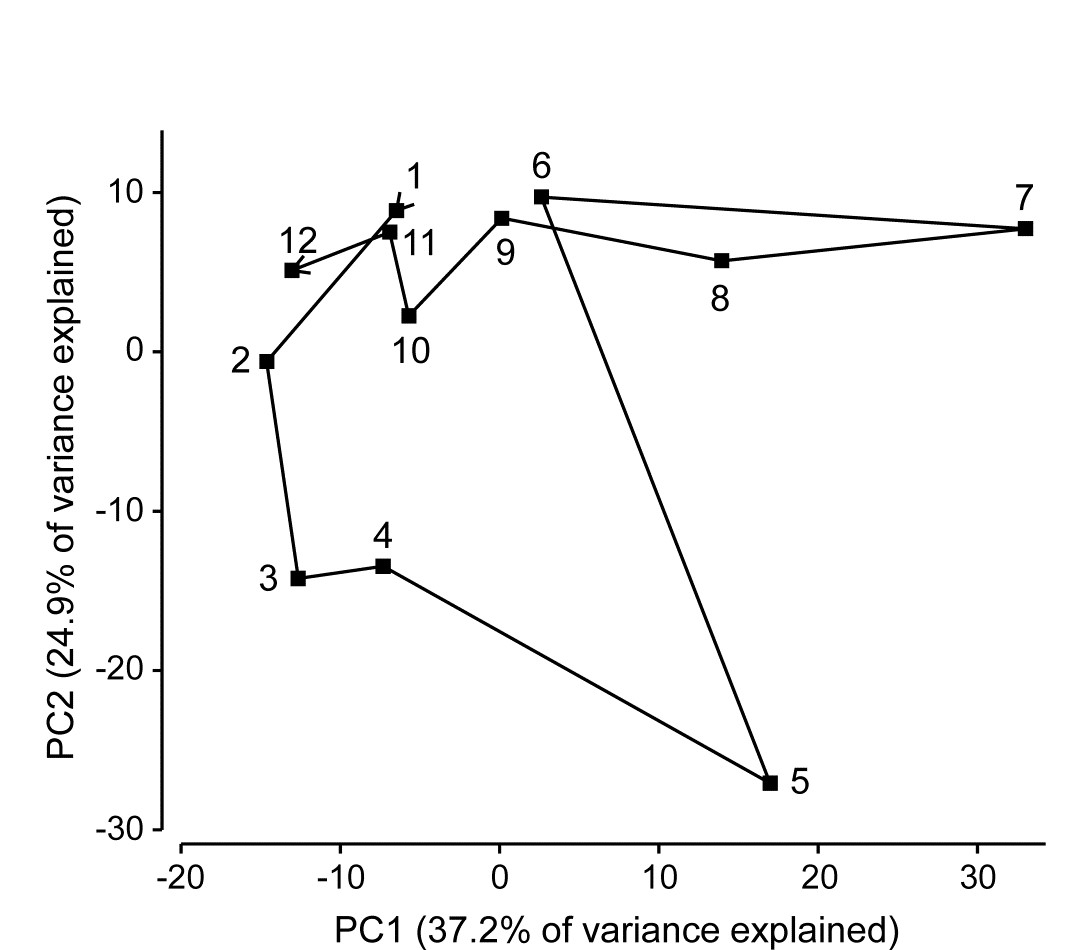
Fig. 4.1. Garroch Head macrofauna {G}. 2-dimensional PCA ordination of square-root transformed biomass of 65 species at 12 sites (1-12) along a transect over the sludge disposal ground at site 6; points joined in transect order (see map in Fig. 1.5).
A total of 11 PC’s are sufficient to capture all the information in this sample matrix, because there are only n=12 samples. (Had n been greater than p, then p PCs would theoretically have been needed to do this, the full PCA then being simply a rotation of the original 65 species axes). However, many fewer than 11 axes are needed to ‘capture’ much of the variability in samples here, the first two axes in Fig. 4.1 explaining 62% of the total variance (a third and fourth would have added another 20% but made no fundamental changes to the broad pattern of this ordination).
There is a puzzling feature to this pattern: the PCA points are joined in their transect order and a natural and interpretable progression of community structure is seen on approach to the dumpsite (1-5) and also on leaving it (7-12). However, site 6 (the dumpsite itself) appears close to sites 1 and 9-12 at the extremes of the transect, suggesting some commonality of the assemblages. Yet examination of the original biomass matrix shows that site 6 has no species in common at all with sites 1 and 9-12! And examination of the environmental data for these sites (on organics, heavy metals and water depth), seen in the later Table 11.1, confirms the expected pattern of contaminant levels in the sediments being greatest at site 6, and least at the transect end-points. The issue here is not that a 2-dimensional PCA is an inadequate description, and that in higher dimensions site 6 would appear well separated from the transect end-points - it does not do so - but that the implicit dissimilarity measure that PCA uses is Euclidean distance, and that is a poor descriptor of differences in biological communities. In other words, the ordination technique itself may not, in some cases, be an inherently defective one, if a high percentage of the original variance is explained in the low-d picture, but the problem is that it starts in the wrong place - with a defective measure of community dissimilarity. The reasons for this are covered in much more detail later, when Euclidean and other resemblance measures are compared for this (and other) data, e.g. Fig. 16.10 on page 16.6.
¶ Later analysis of the count data from this study uses 84 species; 19 of them were too small-bodied to have a weighable biomass.
4.4 PCA for environmental data
The above example makes it clear that PCA is an unsatisfactory ordination method for biological data. However, PCA is a much more useful in the multivariate analysis of environmental rather than species data¶. Here variables are perhaps a mix of physical parameters (grain size, salinity, water depth etc) and chemical contaminants (nutrients, PAHs etc). Patterns in environmental data across samples can be examined in an analogous way to species data, by multivariate ordination, and tools for linking biotic and environmental summaries are fully discussed in Chapter 11.
PCA is more appropriate to environmental variables because of the form of the data: there are no large blocks of zero counts; it is no longer necessary to select a dissimilarity coefficient which ignores joint absences, etc. and Euclidean distance thus makes more sense for abiotic data. However, a crucial difference between species and environmental data is that the latter will usually have a complete mix of measurement scales (salinity in ‰, grain size in $\phi$ units, depth in m, etc). In a multi-dimensional visualisation of environmental data, samples are points referred to environmental axes rather than species axes, but what does it mean now to talk about (Euclidean) distance between two sample points in the environmental variable space? If the units on each axis differ, and have no natural connection with each other, then point A can be made to appear closer to point B than point C, or closer to point C than point B, simply by a change of scale on one of the axes (e.g. measuring PCBs in $\mu$g/g not ng/g). Obviously it would be entirely wrong for the PCA ordination to vary with such arbitrary scale changes. There is one natural solution to this: carry out a correlation-based PCA, i.e. normalise all the variable axes (after transformations, if any) so that they have comparable, dimensionless scales.
The problem does not generally arise for species data, of course, because though a scale change might be made (e.g. from numbers of individuals per core to densities per m2 of sediment surface), the same scale change is made on each axis and the PCA ordination will be unaffected. If PCA is to be used for biotic as well as abiotic analysis, the default position would be to use correlation-based PCA for environmental data and covariance-based PCA for species data (but much better still, use an alternative ordination method such as MDS for species, starting from a more appropriate dissimilarity matrix, such as Bray-Curtis!). For both biotic or abiotic matrices, prior transformation is likely to be beneficial. Different transformations may be desirable for different variables in the abiotic analysis, e.g. contaminant concentrations will often be right-skewed (and require, say, a log transform) but salinity might be left-skewed and need a reverse log transform, see equation (11.2), or no transform at all. The transform issues are returned to in Chapter 9.
PCA strengths
-
PCA is conceptually simple. Whilst the algebraic basis of the PCA algorithm requires a facility with matrix algebra for its understanding, the geometric concepts of a best-fitting plane in the species space, and perpendicular projection of samples onto that plane, are relatively easily grasped. Some of the more recently proposed ordination methods, which either extend or supplant PCA (e.g. Principal Co-ordinates Analysis, Detrended Correspondence Analysis) can be harder to understand for practitioners without a mathematical background.
-
It is computationally straightforward, and thus fast in execution. Software is widely available to carry out the necessary eigenvalue extraction for PCA. Unlike the simplest cluster analysis methods, e.g. the group average UPGMA, which could be accomplished manually in the pre-computer era, the simplest ordination technique, PCA, has always realistically needed computer calculation. But on modern machines it can take small fractions of a second processing time for small to medium sized matrices. Computation time, however, will tend to scale with the number of variables, whereas with MDS, clustering etc, which are based on sample resemblances (and which have lost all knowledge of the species which generated these) computing time tends to scale with (squared) sample numbers.
-
Ordination axes are interpretable. The PC axes are simple linear combinations of the values for each variable, as in equation (4.1), so have good potential for interpretation, e.g. see the Garroch Head environmental data analysis in Chapter 11, Fig. 11.1 and equation (11.1). In fact, PCA is a tool best reserved for abiotic data and this Clyde data set is thus examined in much more detail in Chapter 11.
PCA weaknesses
-
There is little flexibility in defining dissimilarity. An ordination is essentially a technique for converting dissimilarities of community composition between samples into (Euclidean) distances between these samples in a 2- or higher-dimensional ordination plot. Implicitly, PCA defines dissimilarity between two samples as their Euclidean distance apart in the full p-dimensional species space; however, as has been emphasised, this is rather a poor way of defining sample dissimilarity: something like a Bray-Curtis coefficient would be preferred but standard PCA cannot accommodate this. The only flexibility it has is in transforming (and/or normalising) the species axes so that dissimilarity is defined as Euclidean distance on these new scales.
-
Its distance-preserving properties are poor. Having defined dissimilarity as distance in the p-dimensional species space, PCA converts these distances by projection of the samples onto the 2-dimensional ordination plane. This may distort some distances rather badly. Taking the usual visual analogy of a 2-dimensional ordination from three species, it can be seen that samples which are relatively far apart on the PC3 axis can end up being co-incident when projected (perhaps from ‘opposite sides’) onto the (PC1, PC2) plane.
¶ An environmental data matrix can be input to PRIMER in the same way as a species matrix, though it is helpful to identify its Data type as ‘Environmental’ (other choices are ‘Abundance’, ‘Biomass’ or ‘Other’) because PRIMER then offers sensible default options for each type, e.g. in the selection of Resemblance coefficient. In statistics texts, the data matrix is usually described as having n rows (samples) by p columns (variables) whereas the biological matrices we have seen so far have always had species variables as rows (the reason for this convention in biological contexts is clear: p is often larger than n, and binomial species names are much more neatly displayed as row than column labels!). It is not necessary to transpose either matrix type before entry into PRIMER: in the Open dialog, simply select whether the input matrix has samples as rows or columns, or amend that information later (on the Edit>Properties menu) if it has been incorrectly entered initially.
4.5 Example: Dosing experiment, Solbergstrand mesocosm
An example of this final point for a real data set can be seen in Fig. 4.2. This is of nematode data for the dosing experiment {D} in the Solbergstrand mesocosms, at the GEEP Oslo Workshop ( Bayne, Clarke & Gray (1988) ). Box core samples were collected from Oslofjord and held for three months under four dosing regimes: control, low, medium, high doses of a hydrocarbon and Cu contaminant mixture, continuously dosed to the basin waters. Four replicate box cores were subjected to each treatment and at the end of the period cores for all 16 boxes were examined for nematode communities (amongst other faunistic components). Fig. 4.2 shows the resulting PCA, based on log-transformed counts for 26 nematode genera. The interest here, of course, is in whether all replicates from one of the four treatments separate out from other treatments, which might indicate a change in community composition attributable to a directly causal effect of the PAH and Cu contaminant dosing. A cursory glance suggests that the high dose replicates (H) may indeed do this. However, closer study shows that the % of variance explained by the first two PC axes is very low: 22% for PC1 and 15% for PC2. The picture is likely to be very unreliable therefore, and an examination of the third and higher PCs confirms the distortion: some of the H replicates are much further apart in the full species space than this projection into two dimensions would imply. For example, the right-hand H sample is actually closer to the nearest M sample than it is to other H samples. The distances in the full species space are therefore poorly-preserved in the 2-dimensional ordination.
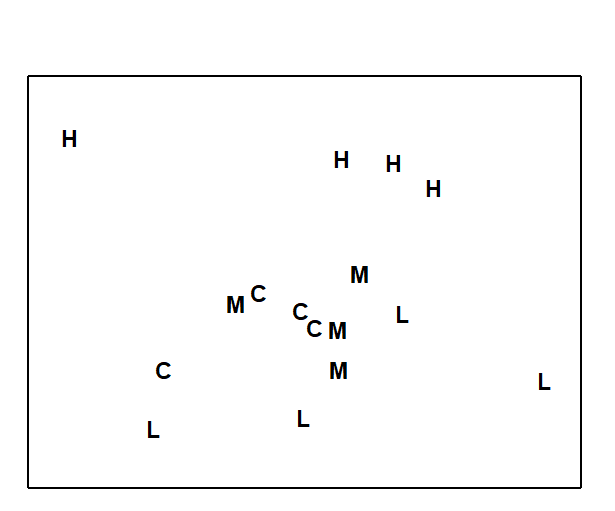
Fig. 4.2. Dosing experiment, Solbergstrand {D}. 2-dimensional PCA ordination of log-transformed nematode abundances from 16 box cores (4 replicates from each of control, low, medium and high doses of a hydrocarbon and Cu contaminant mixture). PC1 and PC2 account for 37% of the total variance.
This example is returned to again in Chapter 5, Fig. 5.5, where it is seen that an MDS of the same data under a more appropriate Bray-Curtis dissimilarity makes a better job of ‘dissimilarity preservation’, though the data is such that no method will find it easy to represent in two dimensions. The moral here is clear:
a) be very wary of interpreting any PCA plot which explains so little of the total variability in the original data;
b) statements about apparent differences in a multivariate community analysis of one site (or time or treatment) from another should be backed-up by appropriate statistical tests; this is the subject of Chapter 6.